
文|聚美丽 诗 诗
在不同的化妆品功效类目中,抗衰老无疑是技术含量最高的一类。目前在市场中,消费者认知度较高的抗衰老产品基本都围绕玻色因、多肽和维A醇三类明星成分为主,它们具备了出色的抗衰功效和完整的实验数据,占据了非常大的市场份额。
对于化妆品企业来说,想要给予消费者有效的抗衰老解决方案,在研究成分和配方技术之外,对皮肤衰老机理这个问题根源的研究也是极为重要的。只有弄明白衰老的“真相”,才能找到抗衰老的有效通路。
衰老是人体的一种自然规律,人们对衰老的研究并不止步于皮肤衰老,还包括病理性衰老,以及外界环境、情绪、生活作息等因素对身体机能老化造成的影响等。事实上,探索衰老的空间很大,在对衰老机理的不断剖析过程中,科学家们从不同的维度出发,提出了各种不同的衰老理论学说。其中比较具有代表性的有:自由基衰老学说、非酶糖基化衰老学说、基因调控学说、线粒体学说等。

此外,与衰老机理相关的理论学说还有很多,虽然来自不同的研究层面,但不同的衰老机理并不是完全对立的,它们能够进行互相补充和验证。

正是因为机理研究的路线错综复杂,关于皮肤衰老的真正原因,至今仍然是一个未解之谜,需要人们持续研究探索。不过,现有的研究结果在相关产品开发中具有重要的指导意义,结合不同的抗衰理论学说,能够为产品研发提供更多思路。
皮肤结构和皮肤衰老过程
在谈及皮肤衰老过程之前,我们先了解皮肤的大致结构。皮肤作为身体和环境之间的保护屏障。它分为三个主要区域:表皮、真皮和皮下组织。
表皮是皮肤的最外层,主要由角质形成细胞和黑素细胞组成。角质形成细胞能够合成角蛋白(即表皮的主要蛋白质结构),并充当激活细胞过程中化学介质的细胞因子。因此,表皮决定了活性物质通过皮肤的迁移深度。
而真皮层主要决定皮肤的紧实程度,拥有最广泛的细胞类型,如神经、血管和淋巴液,周围环绕着由成纤维细胞、胶原蛋白、网状纤维和弹性纤维形成的结缔组织[1]。
此外,表皮和真皮之间还存在一层基底膜,这是一种由生长因子和细胞外基质蛋白(ECM蛋白)富集的结构,主要由表皮细胞和真皮细胞合成和分泌 。值得注意的是,基底膜中的ECM蛋白也提供信号,能够维持表皮细胞处于促增殖状态。
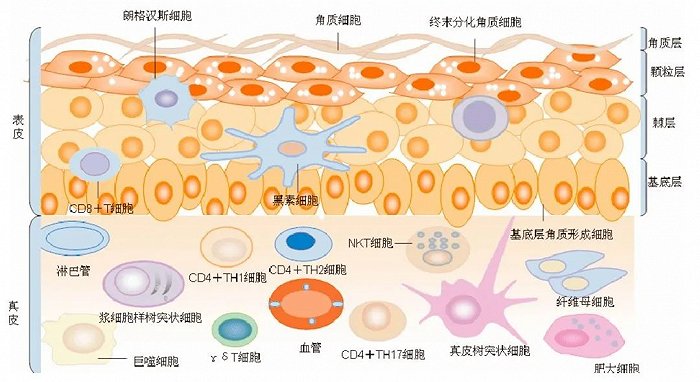
△皮肤结构
衰老是一个复杂且连续的生物过程,其特征是细胞和分子发生变化、身体保持体内平衡的能力逐渐降低、细胞凋亡和衰老的增加。随着时间流逝,这个过程表现得更加明显。
皮肤老化的两个主要过程是内源性老化和外源性老化。内源性老化受个人的遗传控制,随着时间推移逐步显现,这是无法避免的规律;而外源性老化,主要由环境因素引起,比如紫外线辐射、吸烟、风吹、日晒及接触有害化学物质等。
在细胞水平上,皮肤老化也带来了一些功能性的变化,包括角质形成细胞增殖能力下降,角质层形成减少,保护性屏障再生能力下降,脂质合成减少等[2]。
自由基氧化
在现有的十几种衰老机理学说中,自由基学说是具有最大影响力的学说之一。
1956年,美国学者Denham Harman首次提出自由基衰老学说,该学说认为外源和内源产生的自由基,是造成身体的功能下降,导致衰老的主要原因。而维持体内足量的自由基清除剂水平应该可以有效预防衰老、甚至延长寿命。
1969年,美国杜克大学的研究者们发现了体内第一个抗氧化物酶——超氧岐化酶(SOD),并证明SOD作用是体内催化超氧阴离子发生歧化反应产生过氧化氢,这个发现具有非常重大的生物学意义,使人们认识到自由基是机体的的正常成分。
作为细胞代谢的主要参与者,自由基是人体必不可少的一种物质。人体细胞中的线粒体需要通过氧气进行代谢,这个过程中会产生性质十分活泼的自由基。但当自由基过多时,会导致线粒体产生ATP的水平下降、线粒体跨膜电位下降、代谢产物水平及钙离子紊乱,从而产生一系列连锁反应,比如,加速衰老、色素沉着等。
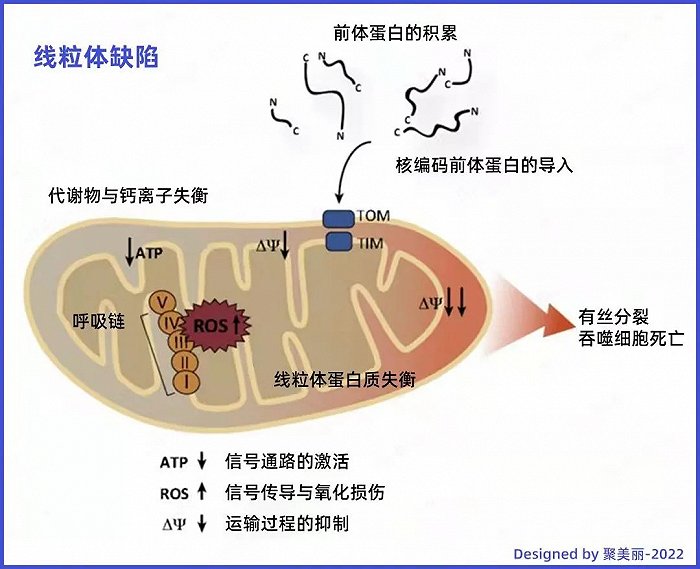
△氧自由基导致线粒体损伤
然而,在后人的研究中,并没有足够有力的实验能够作为支撑抗氧化延缓机体衰老的证据,自由基学说也被认为存在众多缺陷。于是,1990年,美国衰老研究权威Sohal教授首先提出了氧化应激的概念。
氧化应激被定义为氧化和抗氧化之间的平衡失调。活性氧(ROS)是体内或自然环境中由氧组成的活性物质的总称,包括自由基和非自由基(比如过氧化氢、单线态氧分子)。过量的活性氧会导致正常细胞功能所需的生理浓度过高,从而影响细胞代谢[3]。Sohal等人认为,这是造成衰老的根本原因。
细胞的生命活动中不可避免会生成活性氧,其主要有三大来源:巨噬细胞中的NADPH氧化酶、线粒体呼吸链和线粒体多不饱和膜的脂质过氧化。
2016年,发布在《分子科学》上的一篇综述中还提到,活性氧参与皮肤基质形成和降解的信号通路,氧化应激通过降低蛋白酶体活性、损伤DNA及其修复系统、损害端粒等方式影响皮肤衰老。此外,也有研究证明,多环芳烃等污染物、紫外线辐射、吸烟等外在因素,能够加速体内活性氧的生成[4]。
因此,从不同的通道出发,清除体内多余的活性氧和自由基,已经成为目前最常见的抗衰美容方式之一。
皮肤光老化
从皮肤的自然老化历程来看,成年人真皮胶原含量以每年1%的速率递减,胶原的减少主要是基质金属蛋白酶( MMP)表达增加及胶原合成减少所致。随着年龄增长,MMP-1、2、9、12水平升高,而前胶原mRNA表达明显下降,从而导致真皮胶原含量的下降。
与自然老化不同的是,“皮肤光老化”描述了受到外在因素影响,即日光照射下,皮肤的骨架成分胶原蛋白和弹力纤维被紫外线破坏,从而导致皮肤质地发生变化、黑色素增加等。皮肤光老化不仅会影响美观,而且与临床上许多皮肤病密切相关,比如,光化性角化病、光化性弹力纤维病、日光性雀斑样痣、鳞状细胞癌等[5]。
因此,了解光老化的病因及发病机制,有效地对光老化进行预防和治疗已成为目前人们迫切关注的问题。
事实上,关于皮肤光老化的机制理论有很多,其中占据主流的是紫外线致氧自由基(ROS)生成的理论。该理论认为紫外线作用于人体皮肤后,活性氧的生成增多,从而激活丝裂原活化蛋白激酶(MAPK)介导的信号转导途径,进而增强其下游因子激活蛋白-1(AP-1)的表达。
2010年,发表在SCI期刊《光皮肤学,光免疫学与光医学》上的一篇综述中提到,较长波长的紫外光(UVA)穿透较深,可与表皮角质形成细胞和真皮成纤维细胞相互作用,主要通过产生ROS间接发挥作用,随后可产生多种效应,如脂质过氧化、转录因子活化和 DNA 链断裂;而较短波长的紫外光(UVB)则主要影响表皮细胞,即角质形成细胞,其也能够产生ROS,但最主要的作用机制是诱导DNA损伤,与DNA直接作用[6]。
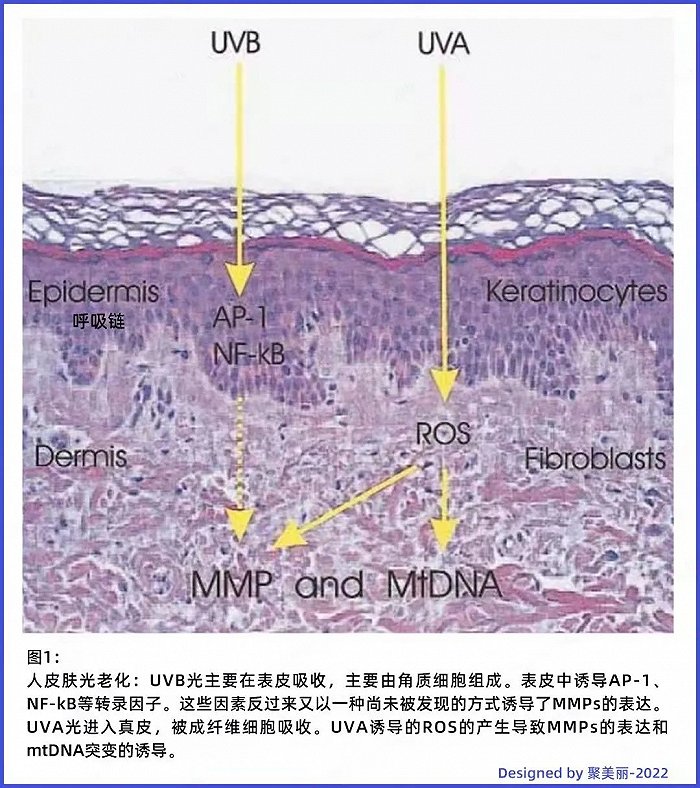
△截图自相关文献
此外,大量证据表明,MMP的诱导在光老化的发病机制中起主要作用。MMP是一组含锌蛋白酶家族,紫外照射可诱导角质形成细胞和成纤维细胞分泌MMPs,继而降解胶原蛋白等真皮细胞外基质成分。相较来说,短波长的UVB能够更快激活MMPs以及诱导DNA损伤。
非酶糖基化
“抗糖化”是最近几年化妆品、食品保健等领域的热门概念,也是抗衰老的重要策略。糖化的全称是“非酶糖基化反应”(NEG),相关的衰老机理研究正成为一个重要方向。
1912年,法国人Louis Camille Maillard最先提出非酶糖基化反应,也称为美拉德反应。非酶糖基化反应首见于食品化学工业领域,是游离还原糖与蛋白质、DNA和脂质的游离氨基发生的非酶促反应,从而产生高级糖基化终产物(AGEs),使蛋白质产生褐色、荧光和交联。
人体内也会发生糖化反应,多余的糖类和蛋白质或脂质作用产生AGEs,此物质的积累会影响细胞稳态和蛋白质结构,与多种皮肤状况有关,包括皮肤的暗沉和衰老。而且,AGEs的形成是不可逆的。
糖基化是随时间自发形成的内源性衰老机制之一。1992年,Yu等人把自由基氧化和非酶糖基化结合起来,提出了非酶糖基化衰老学说[7]。该学说指出:糖基化造成蛋白质的交联损伤是衰老的主要原因。
糖基化还会导致各种蛋白发生交联变性,抗氧化酶和DNA修复酶等功能的损伤;会造成能量供应的减少、代谢功能的降低、平衡失调等老化过程。糖基化终产物AGEs则能够诱导细胞外基质 (ECM) 和细胞损伤,从而导致衰老和与年龄相关的疾病,其存在三种主要机制。
细胞外基质(如胶原蛋白和弹性纤维)内AGEs的积聚,以及AGEs和细胞外基质之间的交联导致结缔组织弹性降低。
细胞内蛋白质的糖基化修饰导致原始细胞功能的丧失。
AGEs 与其晚期糖基化终产物受体 (RAGE) 相互作用,导致激活炎症信号通路、活性氧生成和细胞凋亡[8]。
事实上,形成AGEs的通道主要有三种,包括糖化途径、多元醇途径和糖氧化途径,目前已经发现了约10种AGEs会对皮肤的不同结构有不同程度的影响,它们均能够集中的攻击胶原蛋白。此外,AGEs与RAGE之间的相互作用还会导致机体抗氧化防御减弱、抗氧化能力下降以及炎症的发生[9]。
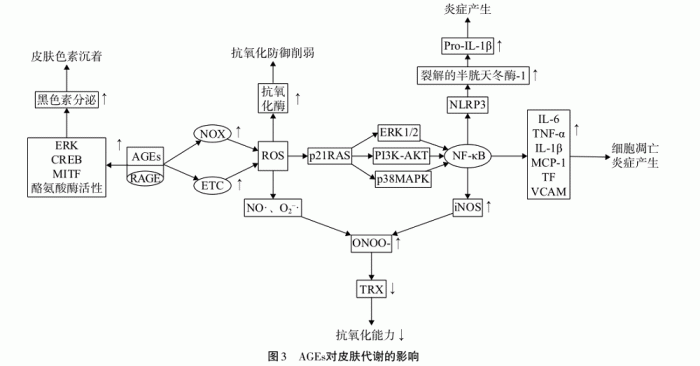
△截图自相关文献
端粒和复制性衰老
现代科学研究表明,端粒和端粒酶与染色体保护、肿瘤发生、细胞衰老等现象密切相关,因而成为科学家当前的研究热点,衰老的端粒学说也在相关机理研究中占据重要地位。
端粒是染色体末端由重复DNA序列和相关蛋白组成的特殊结构,具有稳定染色体结构和完整性的功能;端粒酶是核蛋白逆转录酶,以自身RNA为模板,合成端粒DNA,将端粒DNA加至真核细胞染色体末端,为细胞持续分裂提供遗传基础。
1973年,前苏联科学家Olovnikov提出了端粒学说。该学说认为,在细胞分裂的过程中,端粒会不断变短,直到端粒达到一个临界长度,染色体会失去正常复制的功能,细胞也失去活性而死亡。因此,端粒会随着细胞个体的衰老而变短,端粒损耗的速度称为衡量“生物衰老”的一个指标。
由于端粒缩短,造成DNA损伤,细胞会出现增殖变缓、生长停滞、分化障碍等情况,这种现象称为“复制性衰老”。若细胞中端粒酶活化,端粒长度便可得以维持或延长,从而避免复制老化,阻止内源性衰老进程。
2021年,发表在《Cell》杂志上的一篇文章中提到,端粒几乎与衰老的所有特征都存在内在的相互联系。比如,端粒功能障碍会损害线粒体功能和氧化防御能力,增加ROS水平,构成端粒-p53-PGC1α/β -线粒体的信号传导闭环。此外,端粒功能异常也会影响到组织干细胞耗竭、基因组不稳定、蛋白稳态失衡,甚至会诱导组织炎症反应[10]。
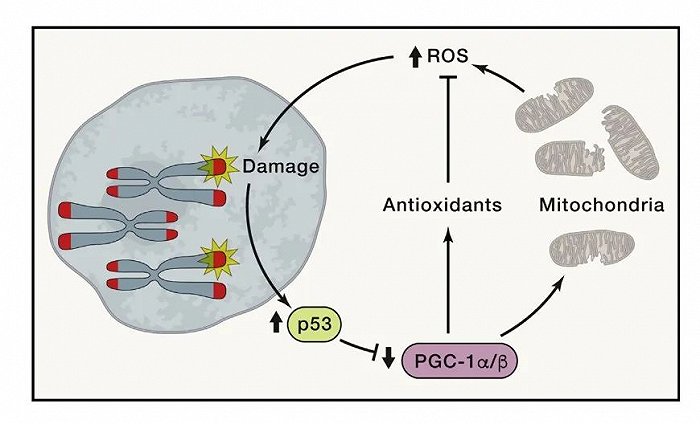
△端粒功能障碍导致线粒体缺陷(截图自相关文献)
2020年,《欧洲分子生物学组织》杂志上发布了一项研究:研究人员从天生端粒过短的先天性角化不良症(dyskeratosis congenita)患者体内收集了大量细胞样本,并进行分析。结果显示,端粒过短会影响到细胞中NAD+(烟酰胺腺嘌呤二核苷酸,俗称抗衰老辅酶Ⅰ)的代谢,导致NAD+不足,进而引发细胞衰老[11]。
此外,在光老化皮肤中,同样存在DNA损伤和端粒缩短的现象,其中可能以端粒损伤为起始反应,再结合UV所致的重要调节基因的共同损伤而加速皮肤的老化。研究表明,紫外线能够加速端粒缩短,同时,光产物环丁烷嘧啶二聚体及和光诱导的活性氧簇分别作用于胸腺嘧啶和鸟嘌呤残基,从而造成端粒袢结构裂解和DNA双链断裂,从而加速老化进程[12]。
虽然端粒从细胞层面解释了内源性衰老的原因,但其只是细胞衰老机理的一个重要组成部分,与细胞、DNA相关的机制研究还有很多方向可以探索。
炎症衰老和免疫系统
“炎症性衰老”是一种与增龄相关的慢性低度炎症的衰老,也是近几年的新兴研究领域。其特点是血液循环中的促炎细胞因子含量增加以及细胞老化的持续变化,这些变化会导致许多增龄相关疾病,其中包括老年痴呆、关节炎、2型糖尿病、与年龄相关的神经退化、皮肤老化等。
纽约大学郎格尼医学中心首席研究员Schneider博士曾说道:“近几十年来,科学界已知炎症、衰老的加速以及癌症是交织在一起的,但它们之间的相互关系基本上是一个谜。”
2014年,《自然通讯》上的一篇研究论文描述了炎症触发细胞衰老,释放包括活性氧簇在内的,与衰老过程相关的一些非常强有力的分子的机制,证明了系统性慢性炎症可通过活性氧介导的端粒功能障碍和细胞衰老而加速衰老[13]。

△截图自相关文献
与此同时,细胞衰老同样会反作用于慢性炎症,从而进一步加剧衰老过程。衰老的成纤维细胞、角质形成细胞会随着衰老过程在皮肤内积聚,并分泌出“衰老相关分泌表型(SASP)”,其中包括促炎细胞因子(如TNF-α、IFN-β、IFN-γ 等)等[14],促炎细胞因子通过产生 ROS 和激活 ATM/ P53/P21( WAF1 / CIP1 )信号通路诱导上皮细胞衰老。
此外,炎症机制可能会加重紫外线的影响,放大对分子和细胞的直接破坏性影响,并影响到免疫系统。
随着年龄增长而出现的免疫功能退化,包括适应性免疫的退化,称为免疫衰老,其特征是因为T细胞激活的中央记忆型T细胞比例降低、效应细胞功能受损、T细胞受体库受限以及抗体多样性的减少,免疫系统做出有效反应的能力降低。
如果免疫系统功能低下或紊乱,则极易导致疾病的发生,包括各种病原微生物的入侵、肿瘤细胞的扩增以及自身免疫病的产生,同时也会导致衰老进程的明显加速。
DNA损伤
2021年4月,德国科隆大学衰老与疾病基因组稳定性研究所Bj rn Schumacher团队在《自然》杂志上发表题为《DNA损伤在衰老过程中的核心作用(The central role of DNA damage in the ageing process)》的综述文章,其详细介绍了DNA损伤与衰老表型的机制联系,并论述了DNA损伤在衰老过程中发挥的核心作用[15]。

△截图自相关文献
核基因组和线粒体基因组会不断受到外源性因素(如紫外线、X射线、食物、水和空气中的化合物)、内源性因素(如活性氧簇、醛类和晚期糖基化终末产物以及自发反应)的破坏。随时间发生的DNA损伤累对分子的影响主要以下有两种:
遗传畸变,如突变和染色体不稳定性;
DNA损伤导致RNA和DNA聚合酶停滞,从而引发DNA损伤信号并干扰主要DNA功能。DNA损伤的细胞和组织后果决定了细胞命运,如细胞死亡和衰老、干细胞衰竭,导致细胞和器官功能丧失、癌症、萎缩和炎症。
文中还提到,核DNA修复造成的NAD+损失、DNA损伤诱导的线粒体自噬缺陷和mtDNA聚合酶表达改变等因素,会导致线粒体功能发生障碍。此外,DNA损伤在分子水平上的影响还包括,基因组不稳定、端粒功能障碍、表观遗传学改变、蛋白应激和线粒体功能受损。
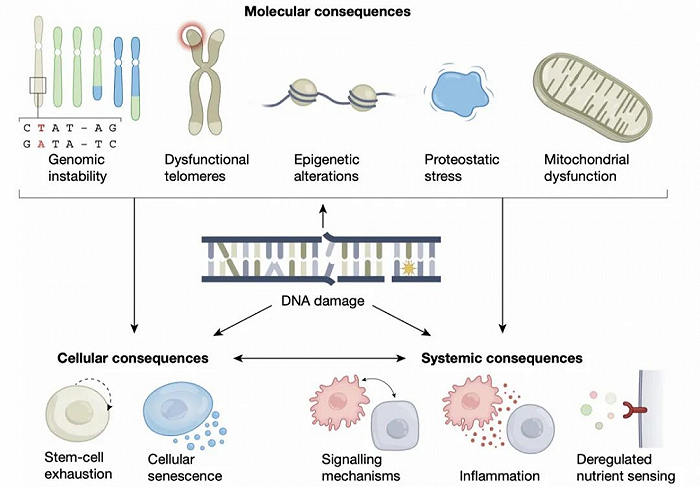
△截图自相关文献
关于机理研究,我们都是在前人的推测基础上,通过实验不断推翻、发展和补充,从而尽可能地接近衰老的真相。尽管对于皮肤衰老的机理研究有很多,但现实是,目前我们仍然未能得知衰老的真正原因。
不过,从研究较为热门的几种衰老机理中,我们能看到,不同的理论学说之间是存在着关联与互通的。对这些衰老机理进行足够了解和跟进,在抗衰老化妆品开发过程中是必不可少的一步。
参考文献:1. R Dearaújo, Lbo M , Trindade K , et al. Fibroblast Growth Factors: A Controlling Mechanism of Skin Aging[J]. Skin pharmacology and physiology, 2019, 32:275-282.
2. DiLoreto R, Murphy CT. The cell biology of aging. Mol Biol Cell. 2015 Dec;26(25):4524–31.
3. 原慧萍, 杨泽. 氧化应激与衰老研究进展[J]. 中国老年保健医学, 2015(5):14-17.
4. Starr J M , Starr R J . Skin Aging and Oxidative Stress[J]. Aging, 2014:15-22.
5. 曾丽, 胡晗菲, 谢红炬,等. 皮肤光老化的研究进展[J]. 中南医学科学杂志, 2015, 43(2):4.
6. Berneburg M H , Plettenberg H , Krutmann J . Photoaging of human skin.[J]. Photodermatology Photoimmunology &Photomedicine, 2010, 16(6):239-244.
7. 来吉祥, 何聪芬, 董银卯. 皮肤衰老机理和抗衰老化妆品的研究进展[C]. 第十一届全国日用化工学术研讨会. 北京日化协会, 2008.
8. Niu Y , Xie T , Ge K , et al. Effects of extracellular matrix glycosylation on proliferation and apoptosis of human dermal fibroblasts via the receptor for advanced glycosylated end products.[J]. Am J Dermatopathol, 2008, 30(4):344-351.
9. 李汇柯, 冯楠, 王闻博,等. 皮肤糖化反应发生机制,影响因素及抗糖化在化妆品行业中的发展现状[J]. 日用化学工业, 51(2):8.
10. Chakravarti D , Labella K A , Depinho R A . Telomeres: history, health, and hallmarks of aging[J]. Cell, 2021, 184(2).
11. Stock A J , et al. Re-quilibration of imbalanced NAD metabolism ameliorates the impact of telomere dysfunction[J]. The EMBO Journal, 2020, 39(21).
12. 高瑛瑛, 骆丹. 皮肤老化和光老化中端粒-端粒酶机制研究进展[J]. 国际皮肤性病学杂志, 2009, 35(1):3.
13. Chronic inflammation induces telomere dysfunction and accelerates ageing in mice[J]. Nature Communications, 2014, 2(4172):4172.
14. https://mp.weixin.qq.com/s/0fzL2f-gIglQZX47cVkurA
15. Schumacher B , Pothof J , Vijg J , et al. The central role of DNA damage in the ageing process[J]. Nature.
视觉设计:乐乐
评论